Акушерство и Гинекология №5 / 2016
Нейробиологические эффекты прогестерона и его метаболитов в акушерской практике
ФГБУ Научный центр акушерства, гинекологии и перинатологии им. академика В.И. Кулакова Минздрава России, Москва;
ГБОУ ВПО Московский государственный медико-стоматологический университет им. А.И. Евдокимова;
ГБОУ ВПО Первый МГМУ им. И.М. Сеченова Минздрава России
Цель исследования. Провести систематический анализ данных, имеющихся в современной литературе, о «не репродуктивных» (нейробиологических) эффектах прогестерона в акушерстве и перинатологии, перспективах заместительной терапии прогестероном у недоношенных новорожденных с неврологическими дефицитами.
Материал и методы. В обзор включены данные зарубежных и отечественных статей, найденных в базе данных Pubmed и Medline по данной теме, опубликованных за последние 10–15 лет.
Результаты. Описана роль основного нейрометаболита прогестерона – аллопрегнанолона в формировании центральной нервной системы плода, миелинизации и ремиелинизации, половой дифференцировке плода. Приведены данные о возможности связи дефицита материнского прогестерона и развития врожденных неврологических заболеваний у новорожденных.
Заключение. Растущий интерес к роли нейростероидов в формировании здорового потомства, экспериментальные и клинические исследования в области нейроэндокринологии открывают перспективы для трансляционной медицины и пересмотра традиционных взглядов на невынашивание беременности и профилактику преждевременных родов, как перинатально-акушерскую проблему.
Стремительное развитие медико-биологических наук и трансляционной медицины расширили наши представления о молекулярных основах жизнедеятельности в различные периоды жизни человека. Особое место при этом занимает изучение биологических «не репродуктивных» эффектов стероидных гормонов, в частности прогестерона, который длительное время рассматривался как эссенциальный гормон – индикатор фертильности менструального цикла и ключевой гормон беременности [1–4]. Интерес исследователей и клиницистов к изучению геномных и внегеномных эффектов прогестерона связан с его участием в метаболических процессах центральной нервной системы (ЦНС), а во время беременности – с влиянием не только на ее успешный исход, но и на ЦНС плода, а также «качество» психического здоровья новорожденного [5, 6]. Настоящий обзор посвящен роли прогестерона в акушерстве и перинатологии.
Метаболизм прогестерона
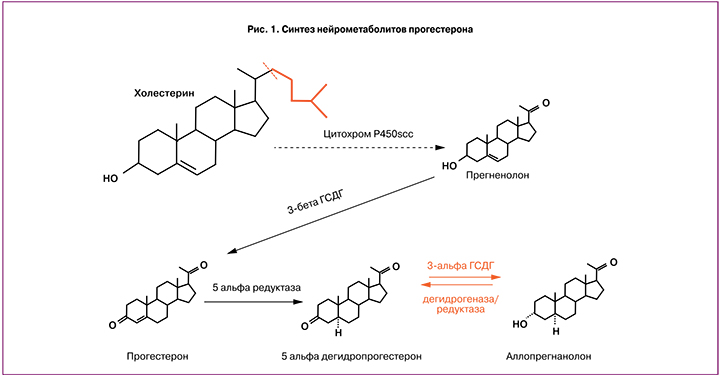
Как и любой стероидный гормон, прогестерон синтезируется из холестерина и формируется в два последовательных этапа: превращение холестерина в прегненолон в митохондриях клеток с помощью фермента цитохрома P450scc, далее под действием 3β-гидроксистероид дегидрогеназы прегненолон трансформируется в прогестерон (рис. 1).
Поскольку прогестерон синтезируется из холестерина, который открывает цепь биосинтеза стероидных половых гормонов и кортикостероидов и содержится повсеместно в клеточных мембранах человека, в том числе в ЦНС, то становятся понятными многогранные эффекты прогестерона на системном, органном, тканевом и локальном уровнях [7].
Классические и неклассические рецепторы прогестерона и его метаболитов
Основной биологический эффект прогестерона осуществляется с помощью классических рецепторов (тип А и тип В), ответственных в основном за геномные эффекты и распространенных во всех отделах головного мозга [5, 8]. Внегеномные эффекты прогестерона осуществляются благодаря его мембранным рецепторам, которые локализуются преимущественно в пре- и постсинаптических отделах нейронов и, предположительно, их взаимодействие с лигандом обеспечивает прямые и быстрые эффекты на синаптическом уровне [9]. Кроме того, данное парасинаптическое расположение экстрагипоталамических рецепторов прогестерона в ЦНС способствует формированию более быстрого ответа на стимул гормона ввиду отсутствия необходимости в прохождении геномного пути реализации соответствующего эффекта [10, 11]. Прогестерон связывается с этими рецепторами с высокой селективностью и сродством, что необходимо для реализации его нейропротекторных свойств [12].
Нейропротекторные свойства прогестерона и аллопрегнанолона
Защитные и трофические эффекты прогестерона и его нейрометаболитов на нервные клетки являются предметом активного изучения. Важнейшим нейрометаболитом прогестерона является аллопрегнанолон, который участвует в метаболических и биохимических процессах головного мозга беременной и развитии нервной системы плода [13]. Важно отметить, что данным метаболитом обладает только натуральный прогестерон. Синтетические гестагены лишены таких производных и, следовательно, не имеют связанных с ними нейропротективных свойств.
В одном из экспериментальных исследований у беременных овец моделировался гипоксический стресс путем пережатия пуповинной артерии. При этом концентрация аллопрегнанолона в головном мозге их плодов значительно повышалась в сравнении с первоначальным уровнем, что свидетельствует о его непосредственном участии в защитных процессах нервной ткани при гипоксических состояниях. При введении ингибитора синтеза аллопрегнанолона уровень гормона в мозге снижался, а количество погибших нервных клеток в результате ишемии значительно увеличивалось. Это позволило предположить, что резкое снижение уровня аллопрегнанолона может способствовать повышенной уязвимости головного мозга к различным видам повреждений [14]. В результате экспериментальных исследований было установлено, что аллопрегнанолон снижал объем зоны ишемии головного мозга у особей как с сохраненной, так и с подавленной экспрессией прогестероновых рецепторов [15, 16].
Психофармакологические свойства аллопрегнанолона (анксиолитическое, антидепрессивное, анестетическое, противосудорожное, болеутоляющее) обусловлены его воздействием на ГАМК (γ-аминомасляная кислота) рецепторы нервных клеток и влиянием на нейрональную возбудимость [17–20].
В последнее время внимание ученых привлекает роль прогестерона и аллопрегнанолона в процессе миелинизации нервных волокон, пролиферации и дифференцировке нервных клеток. Кроме того, аллопрегнанолон участвует в созревании нервных клеток после рождения, способствует дальнейшей миелинизации нервных волокон, что является непрерывным продолжением процесса формирования нервной системы [7]. Миелинизация в ЦНС человека начинается во втором триместре внутриутробного развития. В это время головной мозг плода подвергается влиянию больших концентраций прогестерона и его нейрометаболитов. Уровень циркулирующего прогестерона снижается сразу после рождения и остается на достаточно низком уровне до конца пубертатного периода [21]. Однако в течение первых двух недель жизни концентрация прогестерона преобладает над всеми остальными стероидными гормонами [22]. В это время происходит активный синтез прогестерона и аллопрегнанолона в головном мозге, где наблюдаются их аутокринные и паракринные механизмы действия [7]. При проведении экспериментов на самцах крыс было установлено, что концентрация прогестерона и аллопрегнанолона в головном мозге выше, чем в плазме крови, что может свидетельствовать о непосредственном участии этих гормонов в созревании нервных клеток [16], что крайне важно в формировании ЦНС в целом.
Значение синтеза прогестерона и аллопрегнанолона в головном мозге было изучено на различных животных моделях в неонатальном периоде. Полученные результаты показали, что прогестерон опосредованно через внутриклеточные прогестероновые рецепторы (ПР), а аллопрегнанолон – через ПР-независимые пути (в основном действуя через ГАМК-рецепторы) способствуют созреванию и дифференцировке нейронов, стимулируют формирование дендритов и синаптических контактов между ними, оказывают трофическое влияние на клетки ЦНС [13,16].
Аллопрегнанолон продемонстрировал нейропротекторный эффект в период развития головного мозга в эксперименте на мышах с моделированной болезнью Ниманна–Пика типа C, при которой нарушается внутриклеточный транспорт ганглиозидов холестерина и других липидов, и, соответственно, нарушается синтез стероидных нейрометаболитов [23]. У животных с проведенной заместительной терапией аллопрегнанолоном наблюдалось значительное снижение количества погибших нервных клеток и лучшие неврологические исходы в сравнении с мышами, находившимися без аллопрегнанолоновой гормональной поддержки [24]. Это исследование в очередной раз продемонстрировало важную нейропротекторную роль прогестерона и его метаболитов при развитии ЦНС.

Роль прогестерона и его метаболитов в формировании ЦНС плода
Развитие головного мозга человека идет под постоянным воздействием прогестерона и его метаболитов: 5α-дигидропрогестерона и аллопрегнанолона, которые играют ключевую роль в защите незрелых клеток ЦНС и нервного волокна [19, 21, 25]. Прогестерон и его нейрометаболиты играют фундаментальную роль в формировании и функционировании ЦНС плода [25–27]. Прогестерон стимулирует рост дендритов, соматогенез, спиногенез, а также увеличение конуса роста аксона в незрелых нейронах [28]. Прогестерон и его метаболит аллопрегнанолон снижают вазогенный и цитотоксический отек мозга, оказывают благотворное воздействие на гематоэнцефалический барьер и функции внутричерепного давления, влияют на производство ряда воспалительных цитокинов, увеличивают экспрессию антиоксидантных ферментов, ограничивают глиоз и апоптоз. На фоне применения прогестерона улучшается когнитивное и поведенческое восстановление [29–31]. При этом прогестерон и аллопрегнанолон оказывают эффекты не только через классические ядерные ПР-рецепторы, но и опосредованно через неклассические сигнальные пути (рис. 2).
Действие метаболитов прогестерона на аффективное и социальное поведение осуществляется через ГАМК, глутамат, и дофаминовые рецепторы [27].
Существуют убедительные данные о том, что недостаточный уровень данных стероидов в ЦНС новорожденного при ранних преждевременных родах ведет к нарушению нейрогенеза с задержкой психомоторного и умственного развития, а иногда – к развитию серьезных неврологических заболеваний, в том числе церебрального паралича [25, 26]. В связи с этим в настоящее время активно изучается заместительная прогестероновая поддержка у недоношенных детей. Так, в 2012 году было проведено клиническое исследование с участием младенцев, рожденных до 29 недель беременности с экстремально низкой массой тела, с целью определения влияния прогестероновой заместительной поддержки на неврологические исходы. Одна группа детей получала поддерживающую дозу прогестерона, а другая – плацебо. При достижении детьми 5-летнего возраста исследуемая группа прогестерона показала лучшие когнитивные и неврологические результаты: у них был ниже риск развития церебрального паралича, проявления мышечной спастичности и аметропии [21]. Тем не менее, до сих пор данный вид нейропротекторной гормональной терапии не применяется в практическом здравоохранении. Однако, ввиду особой актуальности проблемы преждевременных родов, не вызывает сомнений необходимость данного направления научного поиска.
Физиологическая роль прогестерона в половой дифференцировке плода
Натуральный прогестерон является субстратом для раннего тестикулярного синтеза тестостерона у плодов мужского пола, который начинается с 6–7-й недели беременности и достигает пика к 12–16-й неделе [32]. Установлена роль прогестерона в половой дифференцировке головного мозга плода. Так, экспериментальные исследования продемонстрировали, что распространенность прогестероновых рецепторов в головном мозге во многом определяет дальнейшую половую дифференцировку [33]. Важно отметить значительную экспрессию прогестероновых рецепторов в ЦНС плодов мужского пола и практически их отсутствие в ЦНС плодов женского пола, что, возможно, определяет дальнейшую дифференцировку сексуального поведения [34].
Влияние нейрометаболитов прогестерона на процессы миелинизации и ремиелинизации
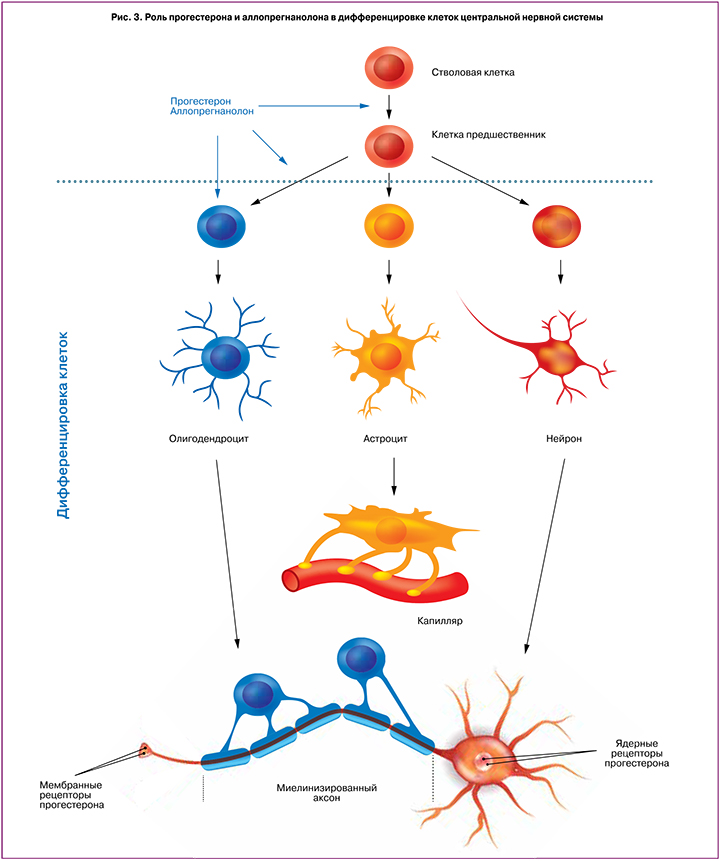
Трудно переоценить роль прогестерона и аллопрегнанолона в процессах миелинизации и формировании белого вещества головного и спинного мозга. В центральной нервной системе аксоны окружаются миелинизированными олигодендроцитами, способствующими быстрому проведению нервного импульса, а также обеспечивающими трофическую функцию нейронов, необходимую для сохранения целостности нейрональной сети. У человека созревание и дифференцировка олигодендроцитов начинается во втором триместре беременности и продолжается длительное время после рождения [25]. Таким образом, формирование белого вещества является довольно длительным и сложным процессом. Во время образования и созревания белое вещество ЦНС довольно чувствительно к повреждающим факторам, особенно в диапазоне от 23 до 32 недель внутриутробного развития. Основной формой повреждения белого вещества головного мозга у недоношенных является перивентрикулярная лейкомаляция, которая лежит в основе развития церебрального паралича [35]. Главными повреждающими факторами при данной патологии являются непосредственно преждевременные роды, внутриутробная инфекция, гипоксические состояния, воспалительные процессы. Во время всего периода развития, когда белое вещество нервной системы наиболее уязвимо, в нем преобладают незрелые олигодендроциты [25, 36]. Повреждающие нейротоксические факторы задерживают развитие и дифференцировку клеток предшественников олигодендроцитов, что ведет к нарушению процесса миелинизации нервных волокон, повреждению белого вещества мозга и развитию неблагоприятных неврологических последствий [25, 37].
Исследование, проведенное на животных моделях, показало важное защитное свойство и прогестерона, и его метаболитов в процессе миелинизации и созревании олигодендроцитов. Данные были получены при изучении гистологических срезов мозжечков мышей. При этом в случае применения прогестерона у мышей наблюдалось умеренное повышение скорости миелинизации нервных волокон [35]. Аллопрегнанолон стимулирует митогенную клеточную активность предшественников олигодендроцитов посредством аутокринных сигнальных механизмов. При этом сами преолигодендроциты синтезируют прогестерон и аллопрегнанолон в значительном количестве, что стимулирует их собственную пролиферацию [36, 37]. На дальнейшем этапе созревания и дифференцировки олигодендроцитов, когда наблюдается наибольшая их уязвимость перед повреждающими нейротоксическими факторами, прогестерон стимулирует пролиферацию и дифференцировку клеток посредством ПР-ассоциированных механизмов (рис. 3).
Однако эти полезные свойства прогестерона могут быть утрачены при действии антагониста ПР (мифепристона). В отличие от этого, аллопрегнанолон, действуя опосредованно через ГАМК-рецепторы, по-прежнему не теряет способность проявлять свои защитные свойства [37]. Важная роль прогестерона при созревании предшественников олигодендроцитов была продемонстрирована в эксперименте при повреждении спинного мозга мышей, когда нарушалась дифференцировка, наблюдалась задержка развития нервных клеток и замедлялся процесс миелинизации в белом веществе [17, 38]. Полученные результаты исследования показали, что полное созревание предшественников олигодендроцитов наблюдалось преимущественно у животных, которые получали поддерживающую гормональную терапию прогестероном [17, 39]. Действие прогестерона, а также его нейрометаболитов было не только направлено на процессы пролиферации преолигодендроцитов, но и оказывало положительное влияние на нейроны, астроглию и нейровоспалительный процесс [36, 40].
Подобные данные получены и при экспериментальном аутоиммунном энцефаломиелите у животных: прогестерон оказывает стимулирующее влияние на процесс миелинизации, обладает нейропротекторным, противовоспалительным действием и улучшает неврологические исходы [41]. В другом исследовании при аутоиммунной демиелинизации нервного волокна под воздействием прогестерона и аллопрегнанолона наблюдались процессы восстановительной ремиелинизации, активное возрождение утраченной миелиновой оболочки [42]. Рядом ученых-исследователей были сделаны выводы, что нарушение синтеза прогестерона и аллопрегнанолона в головном мозге может играть ключевую роль в демиелинизации нервных волокон и приводить к развитию рассеянного склероза [43].
Таким образом, прогестерон, действуя на внутриклеточные ПР, и аллопрегнанолон, действуя на ГАМК-рецепторы как в центральной, так и в периферической нервной системе, способствуют процессам миелинизации и ремиелинизации, воздействуя на Шванновские клетки, которые, как и олигодендроциты в ЦНС, выполняют функцию изоляции аксона, и, как следствие, обеспечивают возможность быстрого сальтаторного проведения нервных импульсов [44]. Несколько проведенных исследований в эксперименте показали важную роль прогестерона и аллопрегнанолона в миелинизации и ремиелинизации после повреждения периферических нервных волокон [42].
Прогестероновый дефицит и неврологические исходы
Этиология большинства врожденных неврологических заболеваний до сих пор остается неизвестной. Одним из таких достаточно распространенных заболеваний является аутизм. Ученые всего мира пытаются найти причину аутизма и разработать эффективные меры его профилактики. В результате проведенного исследования появились небезосновательные предположения, что дефицит прогестерона у матери во время беременности может приводить в дальнейшем к развитию аутизма у детей [45, 46]. Наблюдение было проведено на группе детей, страдающих аутизмом, в сравнении с группой здоровых детей. Полученные результаты свидетельствовали, что вероятность развития аутизма у ребенка, рожденного от матери, имевшей больше факторов риска дефицита прогестерона во время беременности, выше по сравнению с теми, кто не имел этих факторов риска. Авторами была выдвинута теория, что, вероятно, низкая концентрация прогестерона во время беременности приводит к морфологическим изменениям в нервной системе, обусловливающим развитие аутизма [46]. Поэтому очень большое значение имеет поддержание определенного нормального уровня прогестерона в течение всей беременности [47].
Заключение
Приведенные в обзоре данные ставят новые вопросы перед специалистами, осуществляющими наблюдение за течением беременности. Во время внутриутробного развития плод подвергается воздействию материнского прогестерона и его метаболитов, в особенности аллопрегнанолона. Ввиду наличия убедительных доказательств можно с уверенностью констатировать, что эти стероидные гормоны не только играют важную роль в развитии и формировании ЦНС, особенно во второй половине беременности, но и оказывают защитные эффекты при неблагоприятных обстоятельствах, в первую очередь при гипоксических состояниях. Снижение нормальных материнских и, следовательно, фетальных концентраций этих нейростероидов во время беременности может оказывать негативное влияние на развитие головного мозга, нервной системы в целом и приводить к неблагоприятным неврологическим исходам. Поддержание достаточного уровня прогестерона в течение всей беременности играет очень важную роль в развитии нервной системы плода.
Не вызывает сомнения важность применения препаратов натурального прогестерона при прогестерон-дефицитных состояниях в аспекте описанных его нейропротекторных свойств, способности влияния на развитие нервной системы плода. При этом использование синтетических гестагенов не обеспечивает всего этого спектра физиологических нейрометаболических эффектов.
Растущий интерес к роли нейростероидов в формировании здорового потомства, экспериментальные и клинические исследования в области нейроэндокринологии открывают перспективы для трансляционной медицины и пересмотра традиционных взглядов на невынашивание беременности и профилактику преждевременных родов как перинатально-акушерскую проблему.
Список литературы
1. Гончаров Н.П. Кортикостероиды, метаболизм, механизм действия и клиническое применение. М.: Адамантъ; 2002. 169 с.
2. Дедов И.И., ред. Эндокринология. Национальное руководство. М.: Гэотар-Медиа; 2016. 1112 c.
3. Савельева Г.М. Акушерство. М.: Гэотар-Медиа; 2010. 656 c.
4. Якушевская О.В. Современные подходы к назначению прогестерона в практике акушера-гинеколога. Эффективная фармакотерапия. Акушерство и гинекология. 2012; 4: 12-7.
5. Jacobsen B.M., Horwitz K.B. Progesterone receptors, their isoforms and progesterone regulated transcription. Mol. Cell. Endocrinol. 2012; 357(1-2): 18-29.
6. Mani S., Portillo W. Activation of progestin receptors in female reproductive behavior: interactions with neurotransmitters. Front. Neuroendocrinol. 2010; 31(2): 157-71.
7. Schumacher M., Mattern C., Ghoumari A., Oudinet J.P., Liere P., Labombarda F. et al. Revisiting the roles of progesterone and allopregnanolone in the nervous system: Resurgence of the progesterone receptors. Progr. Neurobiol. 2014; 113: 6-39.
8. Brinton R.D., Thompson R.F., Foy M.R., Baudry M., Wang J., Finch C.E. et al. Progesterone receptors: form and function in brain. Front. Neuroendocrinol. 2008; 29(2): 313-39.
9. Waters E.M., Torres-Reveron A., McEwen B.S., Milner T.A. Ultrastructural localization of extranuclear progestin receptors in the rat hippocampal formation. J. Comp. Neurol. 2008; 511(1): 34-46.
10. Peluso J.J., DeCerbo J., Lodde V. Evidence for a genomic mechanism of action for progesterone receptor membrane component-1. Steroids. 2012; 77(10): 1007-12.
11. Thomas P. Characteristics of membrane progestin receptor alpha (mPRalpha) and progesterone membrane receptor component 1 (PGMRC1) and theirroles in mediating rapid progestin actions. Front. Neuroendocrinol. 2008; 29(2): 292-312.
12. Thomas P., Pang Y. Membrane progesterone receptors: evidence for neuroprotective, neurosteroid signaling and neuroendocrine functions in neuronal cells. Neuroendocrinology. 2012; 96(2): 162-71.
13. Brunton P.J., Russell J.A. Allopregnanolone and suppressed hypothalamo-pituitary-adrenal axis stress responses in late pregnancy. Stress. 2011; 14(1): 6-12.
14. Yawno T., Yan E.B., Walker D.W., Hirst J.J. Inhibition of neurosteroid synthesis increases asphyxia-induced brain injury in the late gestation fetal sheep. Neuroscience. 2008; 146(4): 1726-33.
15. Liu A., Margaill I., Zhang S., Labombarda F., Coqueran B., Delespierre B. et al. Progesterone receptors: a key for neuroprotection in experimental stroke. Endocrinology. 2012; 153(8): 3747-57.
16. Zhang Z., Yang R., Cai W., Bai Y., Sokabe M., Chen L. Treatment with progesterone after focal cerebral ischemia suppresses proliferation of progenitor cells but enhances survival of newborn neurons in adult male mice. Neuropharmacology. 2010; 58(6): 930-9.
17. Labombarda F., Ghoumari A.M., Liere P., De Nicola A.F., Schumacher M., Guennoun R. Neuroprotection by steroids after neurotrauma in organotypic spinal cord cultures: a key role for progesterone receptors and steroidal modulators of GABA(A) receptors. Neuropharmacology. 2013; 71: 46-55.
18. Hosie A.M., Wilkins M.E., Smart T.G. Neurosteroid binding sites on GABA(A) receptors. Pharmacol. Ther. 2007; 116: 7-19.
19. Reddy D.S. Neurosteroids: endogenous role in the human brain and therapeutic potentials. Prog. Brain Res. 2010; 186: 113-37.
20. Rupprecht R., Papadopoulos V., Rammes G., Baghai T.C., Fan J., Akula N. et al. Translocator protein (18 kDa) (TSPO) as a therapeutic target for neurological and psychiatric disorders. Nat. Rev. Drug Discov. 2010; 9(12): 971-88.
21. Trotter A., Steinmacher J., Kron M., Pohlandt F. Neurodevelopmental follow-up at five years corrected age of extremely low birth weight infants after postnatal replacement of 17-beta-estradiol and progesterone. J. Clin. Endocrinol. Metab. 2012; 97(3): 1041-7.
22. Robertson N.J., Tan S., Groenendaal F., van Bel F., Juul S.E., Bennet L. et al. Which neuroprotective agents are ready for bench to bedside translation in the newborn infant? J. Pediatr. 2012; 160(4): 544-52.
23. Mellon S.H., Gong W., Schonemann M.D. Endogenous and synthetic neurosteroids in treatment of Niemann-Pick Type C disease. Brain Res. Rev. 2008; 57(2): 410-20.
24. Griffin L.D., Gong W., Verot L., Mellon S.H. Niemann-Pick type C disease involves disrupted neurosteroidogenesis and responds to allopregnanolone. Nat. Med. 2004; 10(7): 704-11.
25. Jakovcevski I., Filipovic R., Mo Z., Rakic S., Zecevic N. Oligodendrocyte development and the onset of myelination in the human fetal brain. Front. Neuroanat. 2009; 3: 5.
26. Lebel C., Beaulieu C. Longitudinal development of human brain wiring continues from childhood into adulthood. J. Neurosci. 2011; 31(30): 10937-47.
27. Frye C.A., Paris J.J., Walf A.A., Rusconi J.C. Effects and mechanisms of 3α,5α,-THP on emotion, motivation, and reward functions involving pregnane xenobiotic receptor. Front. Neurosci. 2012; 5: 136.
28. Theis V., Theiss C. Progesterone: a universal stimulus for neuronal cells? Neural Regen. Res. 2015; 10(4): 547-9.
29. Sayeed I., Stein D.G. Progesterone as a neuroprotective factor in traumatic and ischemic brain injury. Prog. Brain Res. 2009; 175: 219-37.
30. Stein D.G., Wright D.W. Progesterone in the clinical treatment of acute traumatic brain injury. Expert Opin. Investig. Drugs. 2010; 19(7): 847-57.
31. Wright D.W., Yeatts S.D. Very early administration of progesterone for acute traumatic brain injury. N. Engl. J. Med. 2014; 371(26): 2457-66.
32. Стрижаков А.Н., Тимохина Е.В., Игнатко И.В., Белоцерковцева Л.Д. Патофизиология плода и плаценты. М.: ГЭОТАР-Медиа; 2015: 42.
33. Quadros P.S., Pfau J.L. Sex differences in progesterone receptor expression: a potential mechanism for estradiol-mediated sexual differentiation. Endocrinology. 2002; 143(10): 3727-39.
34. Christine K., Ann Nakayama. Potential role of maternal progesterone in the sexual differentiation of the brain. Endocrinology. 1998; 139(8): 3658-61.
35. Shen Y., Plane J.M., Deng W. Mouse models of periventricular leukomalacia. J. Vis. Exp. 2010; May18; (39).
36. Labombarda F., González S.L., Lima A., Roig P., Guennoun R., Schumacher M., de Nicola A.F. Effects of progesterone on oligodendrocyte progenitors, oligodendrocyte transcription factors, and myelin proteins following spinal cord injury. Glia. 2009; 57(8): 884-97.
37. Wang J.M., Liu L., Irwin R.W., Chen S., Brinton R.D. Regenerative potential of allopregnanolone. Brain Res. Rev. 2008; 57(2): 398-409.
38. De Nicola A.F., Labombarda F., Gonzalez Deniselle M.C., Gonzalez S.L., Garay L., Meyer M. et al. Progesterone neuroprotection in traumatic CNS injury and motoneuron degeneration. Front. Neuroendocrinol. 2009; 30(2): 173-87.
39. Labombarda F., González S., Lima A., Roig P., Guennoun R., Schumacher M., De Nicola A.F. Progesterone attenuates astro- and microgliosis and enhances oligodendrocyte differentiation following spinal cord injury. Exp. Neurol. 2011; 231(1): 135-46.
40. Labombarda F., Gonzalez S.L., Gonzalez Deniselle M.C., Vinson G.P., Schumacher M., De Nicola A.F., Guennoun R. Effects of injury and progesterone treatment on progesterone receptor and progesterone binding protein 25-Dx expression in the rat spinal cord. J. Neurochem. 2003; 87(4): 902-13.
41. Garay L., Deniselle M.C., Meyer M., Costa J.J., Lima A., Roig P., De Nicola A. F. Protective effects of progesterone administration on axonal pathology in mice with experimental autoimmune encephalomyelitis. Brain Res. 2009; 1283: 177-85.
42. Hussain R., El-Etr M., Gaci O., Rakotomamonjy J., Macklin W.B., Kumar N. et al. Progesterone and nestorone facilitate axon remyelination: a role for progesterone receptors. Endocrinology. 2011; 152(10): 3820-31.
43. Noorbakhsh F., Ellestad K.K., Maingat F., Warren K.G., Han M.H., Steinman L. et al. Impaired neurosteroid synthesis in multiple sclerosis. Brain. 2011; 134(9): 2703-21.
44. Grenier J., Trousson A., Chauchereau A., Cartaud J., Schumacher M., Massaad C. Differential recruitment of p160 coactivators by glucocorticoid receptor between Schwann cells and astrocytes. Mol. Endocrinol. 2006; 20(2): 254-67.
45. Hill M., Parízek A., Jirásek J.E., Jirkovská M., Dusková M., Klímková M. et al. Is maternal progesterone actually independent of the fetal steroids? Physiol. Res. 2010; 59(2): 211-24.
46. Whitaker-Azmitia P.M., Lobel M., Moyer A. Low maternal progesterone may contribute to both obstetrical complications and autism. Med. Hypotheses. 2014; 82(3): 313-8.
47. Dodds L., Fell D.B., Shea S., Armson B.A., Allen A.C., Bryson S. The role of prenatal, obstetric and neonatal factors in the development of autism. J. Autism Dev. Disord. 2011; 41(7): 891-902.
Поступила 18.03.2016
Принята в печать 25.03.2016
Об авторах / Для корреспонденции
Ходжаева Зульфия Сагдуллаевна, д.м.н., профессор, г.н.с. ФГБУ НЦАГиП им. академика В.И. Кулакова Минздрава России. Адрес: 117997, Россия, Москва, ул. Академика Опарина, д. 4. Телефон: 8 (916) 407-75-67. Е-mail: zkhodjaeva@mail.ruСтеняева Наталья Николаевна, к.м.н., с.н.с. отделения андрологии и урологии ФГБУ НЦАГиП им. академика В.И. Кулакова Минздрава России. Адрес: 117997, Россия, Москва, ул. Академика Опарина, д. 4. Телефон: 8 (903) 538-03-34. Е-mail: nataliasten@mail.ru
Силаев Кирилл Алексеевич, аспирант кафедры акушерства и гинекологии лечебного факультета МГМСУ им. А.И. Евдокимова. Адрес: 127473, Россия, Москва, ул. Делегатская, д. 20, стр. 1. Телефон: 8 (916) 988-16-03. E-mail: silaevkir@gmail.com
Хритинин Дмитрий Федорович, член-корр. РАН, д.м.н., профессор кафедры психиатрии и наркологии лечебного факультета ГБОУ ВПО Первый МГМУ им. И.М. Сеченова Минздрава России. Адрес: 119991, Россия, Москва, ул. Трубецкая, д. 8, стр. 2. Телефон: 8 (495) 609-14-00
Муминова Камилла Тимуровна, резидент ФГБУ НЦАГиП им. академика В.И. Кулакова Минздрава России. Адрес: 117997, Россия, Москва, ул. Академика Опарина, д. 4. Телефон: 8 (916) 373-77-07. Е-mail: kamika91@mail.ru
Для цитирования: Ходжаева З.С., Стеняева Н.Н., Силаев К.А., Муминова К.Т., Хритинин Д.Ф. Нейробиологические эффекты прогестерона и его метаболитов в акушерской практике. Акушерство и гинекология. 2016; 5: 25-32.
http://dx.doi.org/10.18565/aig.2016.5.25-32












