Акушерство и Гинекология №12 / 2014
Биологическая роль молочной кислоты в обеспечении стабильности микроэкосистемы влагалища
1Национальный медико-хирургический центр им. Н.И. Пирогова, Москва, Россия; 2Военно-медицинская академия им. С.М. Кирова Министерства обороны России, Санкт-Петербург
Молочная кислота является важнейшим биологическим субстратом, поддерживающим оптимум биологического гомеостаза микроэкосистемы влагалища, а лекарственнное средство фемилекс в форме вагинальных суппозиториев, содержащих 100 мг молочной кислоты, восстанавливает рН, способствует накоплению гликогена, обладает микробицидным, иммунокоррегирующим свойствами и предназначено для лечения гипо- и анацидных нарушений во влагалищной среде.
С середины 80-х годов прошлого века вопросам различных инфекционных заболеваний вульвы, влагалища, шейки матки стали уделять повышенное внимание. Прежде всего это было связано с ростом частоты цервицитов, вагинитов и главным образом – бактериального вагиноза (БВ), частота которого среди представителей некоторых возрастных групп, жителей городских и сельских поселений, разных профессий женской популяции в нашей стране колебалась в широких пределах: от 16 до 87%. С другой стороны важность изучения данного вопроса определялась теми последствиями, к которым приводили БВ и некоторые другие инфекции нижнего отдела женских половых путей: бесплодию, тяжелым инфекциям после гинекологических операций, родов, невынашиванию беременности и др.
Проведенные ранее фундаментальные исследования [1–4] показали, что содержимое влагалища есть не что иное, как биологическая жидкость строго определенного вида со своими четкими параметрами в норме и при патологии. Впервые было сформулировано, что БВ является дисбактериозом влагалища с совершенно определенным симптомокомплексом, характеризующимся дисбалансом функционирования составных частей микроэкосистемы влагалища.
Микроэкосистема влагалища – это сложная многокомпонентная система, которая включает: 1) анатомическое строение влагалища, шейки матки и промежности; 2) гистологическую структуру вагинального эпителия; 3) биохимические и биофизические параметры влагалищной жидкости (ВЖ); 4) состав микробиоты влагалища с обязательным присутствием протективных ацидофильных микроорганизмов. Целостность и единство функционирования биологического гомеостаза данной системы обеспечивается каскадом взаимодействия нейроэндокринной и иммунной систем, влиянием эндогенных и экзогенных факторов. В результате нарушений на любом уровне микроэкосистемы изменяются биологические свойства влагалищной среды, проявляющиеся угнетением роста и снижением функции лактобактерий и на этом фоне – гиперколонизацией ассоциированной с БВ микрофлорой.
Известно, что клетки поверхностного слоя эпителия влагалища богаты гликогеном, который под действием амилазы ВЖ гидролизуется до декстринов и дисахарида мальтозы. Мальтоза поглощается микроорганизмами и в дальнейшем расщепляется на две молекулы глюкозы. Молочнокислые микроорганизмы утилизируют глюкозу в соответствующих видах брожения.
В физиологических условиях во ВЖ превалирует гомоферментный тип молочнокислого брожения при участии группы Lactobacillus acidophilus, конечным продуктом которого является молочная кислота и в меньшей степени другие кислоты (уксусная, пировиноградная).
В результате каскада ферментативных реакций из одного моля глюкозы образуется два моля АТФ. Энергии 1 моля АТФ достаточно для синтеза 11 г биомассы микроорганизмов. Таким образом, в результате молочнокислого брожения из 1 моля глюкозы образуется энергия, необходимая для синтеза 22 г ацидофильной флоры [3].
 Таким образом, ацидофильные микроорганизмы можно рассматривать как своеобразную ферментативную фабрику, способствующую образованию молочной кислоты в качестве продукта энергообеспечения их генеративной функции.
Таким образом, ацидофильные микроорганизмы можно рассматривать как своеобразную ферментативную фабрику, способствующую образованию молочной кислоты в качестве продукта энергообеспечения их генеративной функции.
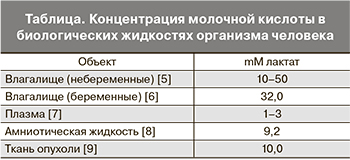
Молочная кислота (МК) – это нормальное промежуточное звено в брожении (окисление, метаболизм) сахара. В природе она образуется в результате молочнокислого брожения и является одним из важнейших метаболитов организма человека – субстратом анаэробного энергетического обеспечения практически всех биореакций. Соли и эфиры МК – лактаты. В организме человека обнаружены D- и L-изомеры МК, которые присутствуют во многих биологических жидкостях и тканях (таблица).
У женщин репродуктивного возраста наибольшее количество МК аккумулируется в просвете влагалища [10, 11, 12]. Она образуется при взаимодействии между слизистой оболочкой влагалища и молочнокислыми бактериями, главным образом лактобактериями. МК во влагалищной жидкости имеет два способа образования: эпителиальное и/или бактериальное. По первому механизму в слое промежуточных вагинальных эпителиальных клеток под влиянием эстрогенов глюкоза метаболизирует при анаэробных условиях до пировиноградной и L-молочной кислот. L-молочная кислота диффузно проникает наружу и скапливается в просвете влагалища.
Второй механизм образования МК заключается в том, что лак...












