Эпидемиология и Инфекционные болезни. Актуальные вопросы №3 / 2012
Крымская геморрагическая лихорадка в Евразии в ХХI веке: эпидемиологические аспекты
ФКУЗ Ставропольский научно-исследовательский противочумный институт Роспотребнадзора; ГОУ ВПО Ставропольская государственная медицинская академия Минздравсоцразвития России; Управление Роспотребнадзора по Ставропольскому краю; Управление Роспотребнадзора по Республике Северная Осетия-Алания, Владикавказ; ФГБУ Институт полиомиелита и вирусных энцефалитов им. М.П. Чумакова РАМН, Москва; ФБУН Центральный НИИ эпидемиологии Роспотребнадзора, Москва
В обзоре рассмотрены молекулярная эпидемиология вируса Крымской-Конго геморрагической лихорадки (ККГЛ), различия и распространенность генотипов вируса ККГЛ, и классическая эпидемиология ККГЛ, клещи-переносчики и хозяева, позвоночные хозяева, пути передачи инфекции человеку и группы риска. Анализ эпидемической ситуации в Евразии и, особенно, в России в XXI веке демонстрирует как рост клинической заболеваемости ККГЛ, так и расширение ареала инфекции на новые географические и экологические зоны. Разработанные и внедренные российскими специалистами меры профилактики ККГЛ могут быть рекомендованы для применения в мировом масштабе и способствовать борьбе с этой тяжелой арбовирусной инфекцией.
Крымская геморрагическая лихорадка (КГЛ) – зоонозная природно-очаговая инфекционная болезнь (код по МКБ-10 – А98.0), вызываемая вирусом Крымской-Конго геморрагической лихорадки (ККГЛ).
Вирусология и молекулярно-генетические аспекты эпидемиологии КГЛ
Вирус ККГЛ принадлежит роду Nairovirus семейства Bunyaviridae. Вирион ККГЛ – сферический, 90–100 нм в диаметре. Геном вируса ККГЛ представляет собой одноцепочечную РНК отрицательной полярности и состоит из 3 сегментов: малого S (кодирует нуклеокапсидный белок N), среднего M (кодирует белок-предшественник, впоследствии расщепляемый SKI-1 и S1P субтилазами и фурин-подобными протеазами на гликопротеины оболочки (Gn и Gc), неструктурный белок GP38 и муцино-подобный белок) и большого L (кодирует полипротеин, включающий РНК-зависимую РНК-полимеразу RdRp). Каждый сегмент имеет только одну транскрипционную единицу, окруженную нетранслируемыми 5’ и 3’ участками, которые необходимы для успешной транскрипции вирусной РНК, формирования вирусных частиц и репликации вируса в целом [1–4]. В конкретный момент времени с РНК вируса ККГЛ осуществляется либо транскрипция мРНК, либо репликация копии генома. Белок N не только участвует в упаковке сегментированной РНК, но и способствует распознаванию ее полимеразой RdRp. Гликопротеин G расщепляется в эндоплазматическом ретикулуме зараженной клетки, затем в виде гетеродимера Gn-Gc связывается с аппаратом Гольджи, в котором накапливаются также рибонуклеопротеиновые комплексы (РНП) [5]. РНП найровирусов производятся в трубкообразных «вирусных фабриках», состоящих из вирусных и клеточных компонентов и соединенных с эндоплазматическим ретикулумом, митохондриями и аппаратом Гольджи. Окончательная сборка вирионов осуществляется в аппарате Гольджи и завершается их экзоцитозом [2, 4].
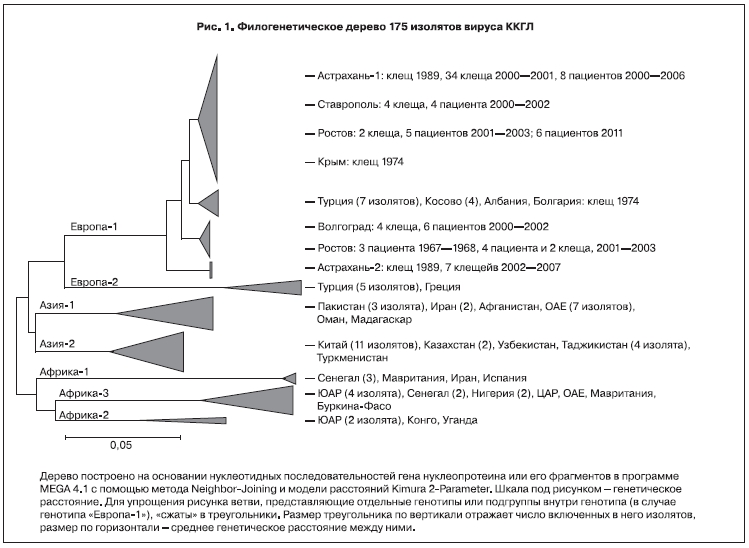
На основании нуклеотидной последовательности S-сегмента выделяют несколько генотипов вируса ККГЛ: генотип I или «Африка-1», II или «Африка-2», III или «Африка-3», IVa или «Азия-1», IVb или «Азия-2», V или «Европа-1», VI или «Европа-2» (рис. 1). «Европа-1» является генотипом, ответственным за 99,9% случаев заболевания КГЛ в России, Турции и на Балканском полуострове, изоляты генотипа «Европа-2» обнаружены только у 3 больных и у 3 экземпляров клещей в Турции в 2007–2008 гг. и у клеща Rhipicephalus bursa в Греции в 1975 г. Генотип «Азия-1» является типичным для Ближнего Востока и юга Азии, генотип «Азия-2» распространен в Китае и среднеазиатских странах СНГ (см. рис. 1). Впрочем, генотип не жестко связан с географическим местом изоляции вируса: разные африканские генотипы выявляются в одном и том же регионе; в Иране и в 2010 г. в Испании выявлены штаммы, принадлежащие генотипу «Африка-1» [6]. Напротив, штаммы, принадлежащие генотипу «Азия-1», изолированы на Мадагаскаре. Генетические деревья, построенные на основании нуклеотидной последовательности L-сегмента, в целом подтверждают существование этих 7 генотипов. Однако анализ имеющихся в настоящее время последовательностей M-сегмента дает несколько иную классификацию. Штаммы «Европа-1» по-прежнему четко выделяются в одну группу (М4), в то время как изоляты генотипов «Африка-3» (типичного для ЮАР), «Азия-1» и часть изолятов «Азия-2» объединяются в группу М2, и напротив, генотип «Азия-2» разбивается на группы М2, М1 и М3 [7].
Характерно, что большинство несинонимичных, ведущих к изменениям аминокислотной последовательности мутаций сосредоточено именно в М-сегменте, особенно в так называемом О-гликозилированном муциноподобном вариабельном участке гликопротеина [8]. Предположительно, частота мутаций в этом участке в 2,3 раза выше, чем в консервативном участке М-сегмента [9]. Изоляты генотипа «Азия-2» наиболее гетерогенны с точки зрения наличия «значимых» аминокислотных замен в нуклеопротеине, то есть замен, меняющих заряд, гидрофобность и плотность упаковки участков аминокислотной последовательности [10]. В целом в популяции изученных штаммов вируса ККГЛ вариабельность по нуклеотидной последовательности для S-, M- и L-сегментов достигает соответственно 20, 31 и 22%; вариабельность по аминокислотной последовательности для этих сегментов составляет соответственно 8, 27 и 10% [11]. Частота мутаций для S-, M- и L-сегментов оценивается приблизительно в 1,1•10-4, 1,5•10-4 и 0,6•10-4 на нуклеотид в год соответственно. Исходя из этого, можно предположить, что общий предок известных генотипов вируса ККГЛ существовал 3000–3500 лет назад [12]. В другой работе [9] авторы дают близкие оценки: 0,3•10-4, 1,2•10-4 и 1,0•10-4 мутаций на нуклеотид в год для S-, M- и L-сегментов, но утверждают, что в генотипе «Африка-3» скорость мутаций выше. Для одноцепочечных вирусов с их несовершенными механизмами репарации вообще характерен активный мутационный процесс, однако у «комариных» арбовирусов обычно скорость мутаций выше, чем у клещевых. Например, у флавивируса Западного Нила скорость мутаций оценивается в 0,3–1...













{kind=link}